Alkuperäiseen artikkeliin nähden olen muuttanut otsikkoa, lisännyt lähdelistan ja muuttanut lauserakenteita ja sanavalintoja hieman. Artikkeli on sisällöltään hieman vaativampaa tasoa kuin useimmat omat kirjoitukseni, mutta luulen että näiden energia-aineenvaihduntaan liittyvien asioiden huolellinen käsittely on kuitenkin ehdottoman tärkeää sairauksien mekanismien ymmärtämiseksi.
Mitokondrioiden toimintahäiriöt ja krooniset sairaudet
1. Mitokondrioiden häiriöt sairauksien mekanismeina
Kroonisia sairauksia on lukuisia. Osa niistä saattaa selittyä samanlaisilla mekanismeilla. Erinomainen esimerkki tästä on vaikkapa autoimmuunisairaudet, joilla on yhteinen autoimmuunimekanismi mutta jotka ilmenevät kuitenkin hyvin erilaisina oirekuvina. Myös mitokondrioiden häiriöt ovat yksi mekanismi joka voi samalla tavalla tuottaa laajan kirjon erilaisia kroonisia sairauksia.
Mitokondrioiden häiriöt ovat potentiaalinen mekanismi useiden aineenvaihdunnallisten, ikääntymiseen liittyvien, neurodegeneratiivisten ja psykiatristen sairauksien taustalla. Esimerkkeinä mainittakoon mm. vanheneminen [1], ikääntymiseen liittyvä verkkokalvon rappeuma [2], Alzheimerin tauti [3], ALS-tauti [4], ateroskleroosi [5], autismi [6], kaksisuuntainen mielialahäiriö [7], syöpä [8], kaihi [2], krooninen väsymysoireyhtymä [9], diabeteksen sydänlihasrappeuma [10], munuaissairaus [11], hermovaurio [12] ja silmäsairaus [2], endoteelin toimintahäiriö [5], epilepsia [13], fibromyalgia [14], kuulonmenetys [15], Huntingtonin tauti [16], verenpainetauti [5], insuliiniresistenssi [17], masennus [7], miehen hedelmättömyys [18], migreeni [19], multippeliskleroosi (?) [20], rasvamaksa [21], lihavuus [22], Parkinsonin tauti [23], skitsofrenia [7] ja tyypin 2 diabetes [24].
Mitokondriohäiriöistä johtuvat sairaudet ja muut elimistön tilat ilmenevät aivoissa, sydämessä, maksassa ja lihaksissa. Mahdollinen syy on se, että kyseiset elimistön osat vaativat paljon aerobista aineenvaihduntaa ja ovat siksi enemmän riippuvaisia mitokondrioista.
“Krooninen mitokondrioiden hapetusstressi perifeerisissä kudoksissa vahingoittaa verkkokalvoa, verisuonten endoteelisoluja, perifeerisiä hermoja ja munuaissoluja, johtaen kliinisesti diabeteksen loppuvaihetta muistuttaviin oireisiin. Täten krooninen mitokondrioiden toimintahäiriö voi selittää kaikki tyypin 2 diabeteksen piirteet, ja on siten sairauden todennäköinen syy."[1]
2. Mitokondriot ja soluhengitys
Mitokondrio on soluissamme sijaitseva soluelin. Endosymbioosi on teoria, jolla selitetään tiettyjen aerobisten bakteerien päätyminen aitotumallisten solujen sisään. Mitokondrioilla on useita bakteerien ominaisuuksia ja omaa DNA:ta (mtDNA).
Mitokondrioiden ensisijainen rooli on tuottaa suurin osa energiastamme aerobisesti soluhengityksellä. Mitokondriot osallistuvat myös detoksifikaatioon sekä apoptoosiin ja energiastatukseen liittyvään solusignalointiin.
Solulimassa glukoosi pilkotaan kuusihiilisestä molekyylisestä kolmihiiliseksi pyruvaatti-molekyyliksi (“palorypälehappo”). Hapettomissa eli anaerobisissa olosuhteissa (esim. intensiivinen liikunta) pyruvaatti muutetaan maitohapoksi*. Tämä prosessi voi tuottaa ATP:tä nopeasti, mutta vain pieniä määriä. Hapellisissa eli aerobisissa olosuhteissa pyruvaatti otetaan mitokondrion sisään ja osallistuu asetyyli-CoA:n muodostumiseen.
Rasvahapot pilkotaan mitokondrioissa, kaksi hiiliatomia kerrallaan, muodostaen näistä palasista asetyyli-CoAa. Asetyyli-CoA vie kaksi hiiliatomia sitruunahappokiertoon, joka poistaa hiilet tuottaen samalla hieman ATP:tä ja sitoen hiilet happeen tuottaen täten hiilidioksidia (CO2). Samalla vetyatomien elektronit sidotaan elektroninkuljettajille, jotka kuljettavat elektronit elektroninsiirtoketjua pitkin tuottaen samalla energiaa protonien pumppaukseen mitokondrion sisäkalvon yli. Kalvon toisella puolella oleva runsas protonien määrä ja toisella puolella oleva runsas elektronien määrä aiheuttaa kalvon yli suuren potentiaalienergia-eron. Entsyymi nimeltä ATP-syntaasi aukaisee kalvolla “portin”, mikä johtaa protonien ja elektronien pariutumiseen ja täten vedyn muodostumiseen. Vety yhdistyy hapen kanssa, jolloin muodostuu vettä. Protonien virratessa ATP-syntaasin läpi vapautuva potentiaalienergia käytetään ATP:n tuotantoon. Suurin osa ATP:stä tuotetaan tätä kautta. (Elektronisiirtoketjun roolia kutsutaan “oksidatiiviseksi fosforylaatioksi” (OxPhos).)
* Eläimimillä ja bakteereilla pyruvaatti käy (fermentoidaan) laktaatiksi eli maitohapoksi. Kasvit ja sienet fermentoivat pyruvaatin etanoliksi.
3. Mitokondriohäiriöiden rooli kroonisissa sairauksissa
Hiilihydraattien ja rasvojen hajottaminen hiilidioksidiksi ja vedeksi ei aina toimi täydellisesti. Joskus veden sijaan muodostuu vetyperoksidia (H2O2), ja elektronin sitoutuminen happeen (O2) (ilmiötä kutsutaan pelkistämiseksi) liian aikaisin voi tuottaa superoksidia (O2-). Vetyperoksidi ja superoksidi ovat yleisiä reaktiivisia happiradikaaleja (ROS). ROS:eilla on rooli solujen signaloinnissa, mutta lisäksi ne varastavat elektroneja (ilmiötä kutsutaan hapettamiseksi) proteiineilta, DNA:lta ja monityydyttymättömiltä rasvahapoilta (lipidiperoksidaatio), vahingoittaen soluja prosessilla jota kutsutaan hapetusstressiksi.
ROS:eilta suojautumista varten meillä on antioksidanttientsyymejä kuten superoksididismutaasi (SOD) (muuttaa superoksidin vetyperoksidiksi), glutationiperoksidaasi ja katalaasi (muuttavat vetyperoksidia vedeksi).
Mikäli ROS:ien määrä ylittää solujen antioksidanttisuojat, ROS:it voivat tuottaa vapaasti hapetusstressiä. Mitokondriot ovat alttiissa asemassa niiden läheisen sijaintinsa vuoksi. ROS:it estävät ATP:n tuotantoa ja voivat vahingoittaa mitokondrioiden DNA:ta aiheuttaen mutaatioita*. Tämä aiheuttaa kierteen, jossa mtDNA-mutaatiot heikentävät mitokondrioiden tuotantoa, hidastaen elektroninsiirtoketjun toimintaa ja lisäten ROS:ien muodostusta, mikä taas johtaa suurempaan määrään hapetusstressiä ja mutaatioita.[1]
Mitokondriot tuottavat noin 90% solujen ROS:eista, joten mitokondrioiden ROS:eilla on paljon potentiaalia aiheuttaa vahinkoa.[1] Vetyperoksidi ja superoksidi ovat vähemmän haitallisia kuin muut hapettimet, mutta ne voivat muodostaa hyvin haitallisia yhdisteitä. Vetyperoksidi voi sitoa metalli-ioneita muodostaen hydroksyyliradikaalin (OH-) ja superoksidi voi yhdistyä typpioksidin (NO) kanssa muodostaen peroksinitriittiä (ONO2–). Mitokondrioiden ROS voi lisätä tulehdussytokiineja[2] ja aiheuttaa insuliiniresistenssiä[3]
Mitokondriot voivat energianpuutteen, hapetusstressin tai kohonneen solunsisäisen kalsiumin (Ca2+) seurauksena käynnistää apoptoosin (ohjelmoitu solukuolema). Nämä kaikki syyt voivat johtua mitokondrioiden toimintahäiriöistä. Apoptoosi ja energianpuute jostakin tietystä tehtävästä vastaavassa soluryhmässä voi johtaa krooniseen sairauteen. Esimerkkeinä mainittakoon haiman beetasolut (tyypin 2 diabetes)**, mustatumakkeen hermosolut (Parkinsonin tauti) ja erilaiset diabetekseen liittyvät ‘-opatiat’ (kardiomyopatia, neuropatia, nefropatia, retinopatia) [1]. Kalsiumionien määrän kasvu voi aiheuttaa eksitotoksisuutta.[4]
Mitokondrioiden toimintahäiriöt ovat vahvasti yhteydessä solulimakalvoston stressiin (ER stress) ja voi myös aiheuttaa sitä ja toisin päin. [5] Solulimakalvosto on soluelin joka osallistuu muun muassa proteiinien synteesiin ja niiden laskostumiseen sekä lipidien ja steroidien synteesiin. Solulimakalvoston stressi johtaa laskostumattomaan proteiinivasteeseen (UPR), joka on adaptiivinen mekanismi, mutta voi pitkään jatkuneena viedä solun apoptoosiin. Pitkään jatkuva solulimakalvoston stressi stimuloi proteiinityrosiinifosfataasi 1B:n (PTP1B) vapautumista, mikä estää leptiini-hormonin signalointia johtaen leptiiniresistenssiin ja lihavuuteen.[6] Lisäksi PTP1B lisää insuliiniresistenssiä, glukoneogeneesiä, triglyseridejä ja kolesterolia [7] sekä lisää syövän kasvua.[8]
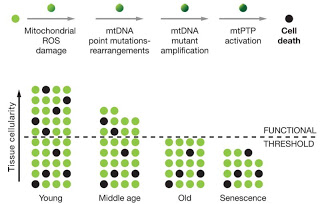
Näiden sairauksien oireet ilmenevät, kun solujen toiminta/määrä laskee tavallisen toimintakyvyn edellyttämän rajan alapuolelle. Mitokondrioiden toimintahäiriö on suuri tekijä monien etenevien, rappeuttavien sairauksien taustalla jotka tyypillisesti ilmenevät ikääntymisen seurauksena. Yleisesti, mitokondrioiden toimintahäiriöt liittyvät monella tavalla myös itse vanhenemiseen ja on käsitykseni mukaan keskeinen tekijä ikääntymisprosessin taustalla.[1]
Olen tehnyt joitakin blogikirjoituksia mitokondriohäiriöiden yhteyksistä lihavuuteen, syöpään, sydän- ja verisuonisairauksiin, tyypin 2 diabetekseen, Parkinsonin tautiin ja Alzheimerin tautiin.
* Toisin kuin tuman DNA:n kohdalla, mitokondrioiden DNA:ta ei voi korjata.[9]
** Tyypin 1 ja 2 diabetekseen liittyy haiman beeta-solujen toiminnan menetys (riittämätön insuliinin eritys). Raskausdiabeteksen kohdalla tilanne on eri, eikä haimassa ole ole vaurioita (myöskään virtsassa ei yleensä havaita sokeria), vaan ensisijainen muutos on suurempi insuliiniresistenssin taso sikiön ravitsemuksen turvaamiseksi. Tämän vuoksi raskausdiabetesta olisi ehkä syytä kutsua mieluummin vain ‘raskausajan insuliiniresistenssiksi’.
*** On lukuisia erilaisia teorioita siitä, mistä vaihdevuodet (menopaussi) johtuvat, joskin olennainen tarkoitus on estää naisten lisääntyminen. Vain äitien mitokondrioiden DNA periytyy lapselle, ja vaihdevuodet koskevat vain naisia. On mahdollista että vaihdevuosien tarkoituksena on estää naisten lisääntyminen siinä vaiheessa kun keho arvioi ikääntymiseen liittyvien mtDNA-mutaatioiden määrän liian suureksi, jotta nämä mutaatiot eivät päätyisi heidän lapsilleen. Tiedetään hyvin, että iäkkäämpien äitien lapsilla on suurempi riski tiettyihin sairauksiin, joten ehkäpä vaihdevuodet ovat vain kehon tapa vetää raja vahinkojen estämiseksi. Vaihdevuodet näyttävät olevan estämätön tapahtuma, joten tämä on vain akateemista spekulaatiota.
4. Mitokondrioiden toimintahäiriöiden syitä
Mitokondrioiden toimintahäiriöitä aiheuttavilla asioilla on tapana aiheuttaa hapetusstressiä. Näihin lukeutuvat mm. alkoholi [1], liikakalorit [2], glukokortikoidit [3], homokysteiini [4], raudan liiallinen kertyminen [5], lipopolysakkaridit (LPS) [6], ravintoainepuutokset [7], tulehdussytokiinit [8], tupakointi [9], ja statiinit* [10].
ATP:n tuotantoa ravintoainekaloreista määrittää tehokkuus (efficiency), johon vaikuttaa mitokondrioiden kytkentä* (coupling). Jos mitokondriot ovat tiukasti kytketty, kalorit muutetaan tehokkaasti ATP:ksi ja heikosti lämmöksi. Liikakalorit lisäävät ROS:ien tuotantoa heikentämällä elektroninsiirtoketjun toimintaa, jolloin elektronit pelkistävät happimolekyylin helpommin superoksidiksi. Mitokondrioiden vähäisempi kytkentä (uncoupling) on kylmässä ilmastossa ilmenevä adaptaatio, jossa kaloreista muodostetaan vähemmän ATP:tä mutta enemmän lämpöä (termogeneesi) eivätkä liikakalorit tuota happiradikaaleja (ROS) yhtä paljoa. Niillä ihmisillä, joilla mitokondriot ovat vahvemmin kytketty, kasvaa liikakaloreista johtuvien mitokondrioiden häiriöiden riski.[2]
Tiettyjen ravinteiden puutokset (mm. CoQ10 [10], glutationi [11], karnitiini [12], tauriini [13] tai sinkki [14]) voivat johtaa mitokondrioiden häiriöihin. Myös muiden mitokondrioiden toimintaa suojaaiven ravinteiden puute voi lisätä hapetusstressiä ja häiritä mitokondrioiden toimintaa. Näihin ravinteisiin lukeutuvat muun muassa:
- Glutationi ja seleeni, joista tuotetaan glutationiperoksidaasia (GPx)
- Mangaani, jota tarvitaan superoksididismutaasin tuotantoon mitokondrioissa (sinkki ja kupari auttavat tuottamaan superoksididismutaasia muissa kehon osissa).
- Vitamiinit B2, B5, B6 ja B7 sekä rauta, kupari ja sinkki tuottavat hemiä mitokondrioissa. Hemi on katalaasin ja mitokondrion IV-kompleksin osa, vähentäen ROSia.[15]
- FAD+ riboflaviinista (B2), NAD+ niasiinista (B3) ja CoQ10 ovat elektroninsiirtoketjun elektroninkuljettajia. Lisäksi, asetyyli-CoA tuotetaan B5-vitamiinista.
- CoQ10 [10], PQQ [16], lipoiinihappo [17], karnitiini [18] karnosiini [19], kreatiini [20], tauriini [21] sekä C- ja E-vitamiinit ovat mitokondrioantioksidantteja, ja niillä saattaa olla myös muita käyttötarkoituksia.
Voit katsoa tästä kirjoituksestani mistä mainittuja ravintoaineita saadaan. Yleisesti eläinperäiset ruoat ja kasvikset ovat parhaita lähteitä. Aminohappojohdannaisia ja CoQ10:ä löytyy lähes ainoastaan lihatuotteista, mikä on ymmärrettävää sillä lihakset ja sisäelimet sisältävät paljon mitokondrioita.
* Kolesterolin alentamiseksi statiinit estävät myös CoQ10-synteesiä. CoQ10-puutos voi aiheuttaa mitokondrioiden toimintahäiriön, mikä voi selittää joitakin statiinien sivuvaikutuksia (esim. kohonnut diabeteksen tai myopatian riski).
** Tiukasti kytketyt mitokondriot ovat tehokkaampia proteiinien pumppaamisessa sisemmälle mitokondriokalvolle, ja tehokkaampia näiden protonien hyödyntämisessä ATP:n tuotannossa, kun ne virtaavat ATP-syntaasin läpi.[2]
*** Ympäristömyrkyt kuten dioksiinit [22], raskasmetallit (alumiini [23] ja elohopea [24]), rikkaruohomyrkyt [25], tuholaismyrkyt [26] ja hitsaushöyryt [27] saattavat myös vaikuttaa mitokondriohäiriöiden syntyyn.
5. Mitokondrioiden biogeneesi
Jos mitokondrioiden lukumäärä ja/tai toiminta on alhainen, silloin olennainen tapa korjata tilanne on lisätä mitokondrioiden biogeneesiä eli uusien mitokondrioiden kasvua. AMP:n aktivoima proteiinikinaasi (AMPK) is energiasensori jota vapautuu solun matalan energiatason seurauksena (korkea AMP:ATP -suhde). Lisäksi AMPK lisää mitofagiaa (mitokondrioiden autofagiaa)[1], glukoosin kuljetusta ja rasvahappojen hapettamista.[2] AMPK lisää PGC-1α:ta, joka on mitokondrioiden biogeneesistä vastuussa oleva proteiini. PGC-1α lisää myös rasvahappojen hapettamista, oksidatiivista fosforylaatiota (OxPhos), insuliiniherkkyyttä sekä energiankulutusta.[3]
Liikunta* [4], ketogeeniset ruokavaliot [5], paastoaminen [6], kalorirajoitus [7], kylmäaltistus [8] ja ei-ketogeeniset vähähiilihydraattiset ruokavaliot [9] lisäävät AMPK:ta. Kaikki näistä viestivät energian vähäisyyttä, joten ovat jossain määrin stressaavia elimistölle ja voivat suurina määrinä heikentää kilpirauhasen toimintaa.
Aktiivinen kilpirauhashormoni (T3) [10], haaraketjuiset aminohapot (BCAA) [11], butyraatti [12], PQQ [13], sekä lipoiinihapon ja karnitiinin yhdistelmä [14] lisäävät PGC-1α:ta (mahdollisesti AMPK:sta itsenäisellä mekanismilla?). Tämä lista on koostettu yleisesti terveyttä tukevista molekyyleistä.
* Mark’s Daily Apple -artikkelin tutkimusten pohjalta: kestävyysurheilu ja HIIT (30sec juoksua-4min taukoa, neljästi) lisää mitokondrioiden biogeneesiä (MB). Lihasharjoittelu lisää kestävyysurheilun aiheuttamaa MB:tä, mutta lihasharjoittelu tai kävely eivät riitä. Veikkaisin että liikunta joka lisää hengittämistä eniten, lisää myös MB:tä eniten. Korkeat annokset C-vitamiinia (1000mg päivässä) vähentävät liikunnan aiheuttamaa MB:tä.[14]
6. Yhteenveto
Mitokondrioiden puutteellinen toiminta on monien aineenvaihdunnallisten, ikääntymiseen liittyvien, neurodegeneratiivisten ja psykiatristen sairauksien taustalla oleva mekanismi. Sen mahdollisia aiheuttajia ovat liikakalorit, ravintoainepuutokset ja hapetusstressi.
Mitokondrioiden toimintahäiriö on kierre, jossa hapetusstressi vahingoittaa mitokondrion DNA:ta heikentäen mitokondrioiden toimintaa ja lisäten hapetusstressiä entisestään.
Monia mitokondrioita tukevia ravintoaineita löytyy eläinkunnan ruoista ja vihanneksista. Näitä ravinteita runsaasti sisältävä ruokavalio sekä mitokondrioiden biogeneesiä lisäävät ja hapetusstressiä vähentävät toimenpiteet voivat auttaa estämään, ellei jopa kumoamaan, mitokondrioiden toimintahäiriön ja muita siihen liittyviä sairauksia.
Lähteet
Osa 1: Mitokondrioiden puutteellinen toiminta sairauksien mekanismina
[1] Annu Rev Genet. 2005;39:359-407. A mitochondrial paradigm of metabolic and degenerative diseases, aging, and cancer: a dawn for evolutionary medicine. Wallace DC.
[2] Ophthalmic Res. 2010;44(3):179-90. The importance of mitochondria in age-related and inherited eye disorders. Jarrett SG, Lewin AS, Boulton ME.
[3] Int J Alzheimers Dis. 2011 Mar 15;2011:925050. Amyloid-Beta interaction with mitochondria. Pagani L, Eckert A.
[4] Pizzuti, A., & Petrucci, S. (2011). Mitochondrial disfunction as a cause of ALS. Archives italiennes de biologie, 149(1), 113-119.
[5] Circ Res. 2007 Mar 2;100(4):460-73. Mitochondrial dysfunction in atherosclerosis. Madamanchi NR, Runge MS.
[6] PLoS One. 2008;3(11):e3815. Mitochondrial disease in autism spectrum disorder patients: a cohort analysis. Weissman JR, Kelley RI, Bauman ML, Cohen BH, Murray KF, Mitchell RL, Kern RL, Natowicz MR.
[7] Chang Gung Med J. 2009 Jul-Aug;32(4):370-9. Mitochondrial dysfunction and psychiatric disorders. Jou SH, Chiu NY, Liu CS.
[8] Cell Cycle. 2011 Dec 1;10(23):4065-73. Mitochondrial oxidative stress in cancer-associated fibroblasts drives lactate production, promoting breast cancer tumor growth: understanding the aging and cancer connection. Balliet RM, Capparelli C, Guido C, Pestell TG, Martinez-Outschoorn UE, Lin Z, Whitaker-Menezes D, Chiavarina B, Pestell RG, Howell A, Sotgia F, Lisanti MP.
[9] Int J Clin Exp Med. 2009;2(1):1-16. Chronic fatigue syndrome and mitochondrial dysfunction. Myhill S, Booth NE, McLaren-Howard J.
[10] Biochim Biophys Acta. 2011 Jul;1813(7):1351-9. Mitochondrial dysfunction in diabetic cardiomyopathy. Duncan JG.
[11] Curr Med Chem. 2010;17(34):4256-69. Oxidative stress in diabetic nephropathy. Kashihara N, Haruna Y, Kondeti VK, Kanwar YS.
[12] Expert Rev Endocrinol Metab. 2010 Jan 1;5(1):39-49. Mitochondrial stress and the pathogenesis of diabetic neuropathy. Fernyhough P, Roy Chowdhury SK, Schmidt RE.
[13] J Bioenerg Biomembr. 2010 Dec;42(6):449-55. Mitochondrial dysfunction and oxidative stress: a contributing link to acquired epilepsy? Waldbaum S, Patel M.
[14] Cordero, M. D., de Miguel, M., & Alcázar, J. A. S. The Role of Oxidative Stress and Mitochondrial Dysfunction in the Pathogenesis of Fibromyalgia.
[15] Cell Res. 2007 Mar;17(3):227-39. Mitochondrial rRNA and tRNA and hearing function. Xing G, Chen Z, Cao X.
[16] Hum Mol Genet. 2010 Oct 15;19(20):3919-35. Mitochondrial loss, dysfunction and altered dynamics in Huntington's disease. Kim J, Moody JP, Edgerly CK, Bordiuk OL, Cormier K, Smith K, Beal MF, Ferrante RJ.
[17] Circ Res. 2008 Feb 29;102(4):401-14. Role of mitochondrial dysfunction in insulin resistance. Kim JA, Wei Y, Sowers JR.
[18] Hum Reprod Update. 2009 Sep-Oct;15(5):553-72. Mitochondrial functionality in reproduction: from gonads and gametes to embryos and embryonic stem cells. Ramalho-Santos J1, Varum S, Amaral S, Mota PC, Sousa AP, Amaral A.
[19] Rev Hosp Clin Fac Med Sao Paulo. 2002 Nov-Dec;57(6):293-8. New migraine preventive options: an update with pathophysiological considerations. Bigal ME, Rapoport AM, Sheftell FD, Tepper SJ.
[20] Neuropathol Appl Neurobiol. 2008 Dec;34(6):577-89. Review: Mitochondria and disease progression in multiple sclerosis. Mahad D, Lassmann H, Turnbull D.
[21] World J Gastroenterol. 2008 Jan 14;14(2):193-9. Nonalcoholic fatty liver disease and mitochondrial dysfunction. Wei Y, Rector RS, Thyfault JP, Ibdah JA.
[22] Steven Hamley - Leptin Resistance
[23] Mol Neurobiol. 2011 Apr;43(2):80-6. Mitochondrial quality control and Parkinson's disease: a pathway unfolds. de Castro IP, Martins LM, Loh SH.
[24] Exp Diabetes Res. 2012;2012:703538. Mitochondrial dysfunction and β-cell failure in type 2 diabetes mellitus. Ma ZA, Zhao Z, Turk J.
Osa 3: Mitokondrioiden toimintahäiriöiden rooli sairauksissa
[1] Annu Rev Genet. 2005;39:359-407. A mitochondrial paradigm of metabolic and degenerative diseases, aging, and cancer: a dawn for evolutionary medicine. Wallace DC.
[2] J Exp Med. 2011 Mar 14;208(3):417-20. Mitochondrial reactive oxygen species drive proinflammatory cytokine production. Naik E, Dixit VM.
[3] Circ Res. 2008 Feb 29;102(4):401-14. Role of mitochondrial dysfunction in insulin resistance. Kim JA, Wei Y, Sowers JR.
[4] J Neurosci. 1996 Oct 1;16(19):6125-33. Mitochondrial dysfunction is a primary event in glutamate neurotoxicity. Schinder AF, Olson EC, Spitzer NC, Montal M.
[2] J Exp Med. 2011 Mar 14;208(3):417-20. Mitochondrial reactive oxygen species drive proinflammatory cytokine production. Naik E, Dixit VM.
[3] Circ Res. 2008 Feb 29;102(4):401-14. Role of mitochondrial dysfunction in insulin resistance. Kim JA, Wei Y, Sowers JR.
[4] J Neurosci. 1996 Oct 1;16(19):6125-33. Mitochondrial dysfunction is a primary event in glutamate neurotoxicity. Schinder AF, Olson EC, Spitzer NC, Montal M.
[5] Exp Diabetes Res. 2012;2012:242984. Interaction between mitochondria and the endoplasmic reticulum: implications for the pathogenesis of type 2 diabetes mellitus. Leem J, Koh EH.
[6] Mol Pharmacol. 2008 Dec;74(6):1610-9. Endoplasmic reticulum stress induces leptin resistance. Hosoi T, Sasaki M, Miyahara T, Hashimoto C, Matsuo S, Yoshii M, Ozawa K.
[7] J Clin Metab Diabetes. 2010 May;1(1):27-33. Hepatic PTP1B Deficiency: The Promise of a Treatment for Metabolic Syndrome? Bence KK.
[8] Trends Biochem Sci. 2010 Aug;35(8):442-9. PTP1B: a double agent in metabolism and oncogenesis. Yip SC, Saha S, Chernoff J.
[9] Cleve Clin J Med. 2001 Jul;68(7):625-6, 629-42. Mitochondrial cytopathy in adults: what we know so far. Cohen BH, Gold DR.
Osa 4: Mitokondrioiden toimintahäiriöiden syitä
[1] Gastroenterology. 2002 Jun;122(7):2049-63. Alcohol and mitochondria: a dysfunctional relationship. Hoek JB, Cahill A, Pastorino JG.
[2] Annu Rev Genet. 2005;39:359-407. A mitochondrial paradigm of metabolic and degenerative diseases, aging, and cancer: a dawn for evolutionary medicine. Wallace DC.
[3] Victor Mark Tang: Glucocorticoid effects on oxidative stress and mitochondrial dysfunction (MSc thesis @ The University Of British Columbia)
[4] J Neurosci. 2000 Sep 15;20(18):6920-6. Homocysteine elicits a DNA damage response in neurons that promotes apoptosis and hypersensitivity to excitotoxicity. Kruman II, Culmsee C, Chan SL, Kruman Y, Guo Z, Penix L, Mattson MP.
[13] Exp Clin Cardiol. 2011 Winter;16(4):125-8. Role of mitochondrial permeability transition in taurine deficiency-induced apoptosis. Jong CJ, Azuma J, Schaffer SW.
[14] J Nutr. 2003 Aug;133(8):2543-8. Zinc deficiency induces oxidative DNA damage and increases p53 expression in human lung fibroblasts. Ho E, Courtemanche C, Ames BN.
[15] = [7]
[16] J Nutr. 2006 Feb;136(2):390-6. Pyrroloquinoline quinone modulates mitochondrial quantity and function in mice. Stites T, Storms D, Bauerly K, Mah J, Harris C, Fascetti A, Rogers Q, Tchaparian E, Satre M, Rucker RB.
[1] Cell Metab. 2011 Feb 2;13(2):119-20. AMPK-dependent phosphorylation of ULK1 induces autophagy. Zhao M, Klionsky DJ.
[2] Proc Natl Acad Sci U S A. 2002 Dec 10;99(25):15983-7. AMP kinase is required for mitochondrial biogenesis in skeletal muscle in response to chronic energy deprivation. Zong H, Ren JM, Young LH, Pypaert M, Mu J, Birnbaum MJ, Shulman GI.
[3] Adv Physiol Educ. 2006 Dec;30(4):145-51. PGC-1alpha: a key regulator of energy metabolism. Liang H, Ward WF.
[4] Biochem J. 2009 Mar 1;418(2):261-75. AMPK and the biochemistry of exercise: implications for human health and disease. Richter EA, Ruderman NB.
[3] Victor Mark Tang: Glucocorticoid effects on oxidative stress and mitochondrial dysfunction (MSc thesis @ The University Of British Columbia)
[4] J Neurosci. 2000 Sep 15;20(18):6920-6. Homocysteine elicits a DNA damage response in neurons that promotes apoptosis and hypersensitivity to excitotoxicity. Kruman II, Culmsee C, Chan SL, Kruman Y, Guo Z, Penix L, Mattson MP.
[5] Proc Natl Acad Sci U S A. 2002 Feb 19;99(4):2264-9. Iron deficiency and iron excess damage mitochondria and mitochondrial DNA in rats. Walter PB, Knutson MD, Paler-Martinez A, Lee S, Xu Y, Viteri FE, Ames BN.
[6] Free Radic Biol Med. 2011 May 15;50(10):1336-43. Mitochondrial dysfunction promoted by Porphyromonas gingivalis lipopolysaccharide as a possible link between cardiovascular disease and periodontitis. Bullon P, Cordero MD, Quiles JL, Morillo JM, del Carmen Ramirez-Tortosa M, Battino M.
[7] Proc Natl Acad Sci U S A. 2006 Nov 21;103(47):17589-94. Low micronutrient intake may accelerate the degenerative diseases of aging through allocation of scarce micronutrients by triage. Ames BN.
[8] Free Radic Biol Med. 2009 Feb 15;46(4):462-70. TNF-induced mitochondrial damage: a link between mitochondrial complex I activity and left ventricular dysfunction. Mariappan N, Elks CM, Fink B, Francis J.
[9] Circ Res. 2007 Mar 2;100(4):460-73. Mitochondrial dysfunction in atherosclerosis. Madamanchi NR, Runge MS.
[10] Ochsner J. 2010 Spring;10(1):16-21. Coenzyme q10 and statin-induced mitochondrial dysfunction. Deichmann R, Lavie C, Andrews S.
[11] Proc Natl Acad Sci U S A. 1991 Mar 1;88(5):1913-7. Glutathione deficiency leads to mitochondrial damage in brain. Jain A, Mårtensson J, Stole E, Auld PA, Meister A.
[12] J Biol Chem. 2009 Aug 21;284(34):22840-52. Carnitine insufficiency caused by aging and overnutrition compromises mitochondrial performance and metabolic control. Noland RC, Koves TR, Seiler SE, Lum H, Lust RM, Ilkayeva O, Stevens RD, Hegardt FG, Muoio DM.
[13] Exp Clin Cardiol. 2011 Winter;16(4):125-8. Role of mitochondrial permeability transition in taurine deficiency-induced apoptosis. Jong CJ, Azuma J, Schaffer SW.
[14] J Nutr. 2003 Aug;133(8):2543-8. Zinc deficiency induces oxidative DNA damage and increases p53 expression in human lung fibroblasts. Ho E, Courtemanche C, Ames BN.
[15] = [7]
[16] J Nutr. 2006 Feb;136(2):390-6. Pyrroloquinoline quinone modulates mitochondrial quantity and function in mice. Stites T, Storms D, Bauerly K, Mah J, Harris C, Fascetti A, Rogers Q, Tchaparian E, Satre M, Rucker RB.
[17] Pharmacol Rep. 2011;63(4):849-58. Lipoic acid - biological activity and therapeutic potential. Gorąca A, Huk-Kolega H, Piechota A, Kleniewska P, Ciejka E, Skibska B.
[18] Drug Discov Today Dis Mech. 2009;6(1-4):e31-e39. CARNITINE HOMEOSTASIS, MITOCHONDRIAL FUNCTION, AND CARDIOVASCULAR DISEASE. Sharma S, Black SM
[19] PLoS One. 2011 Mar 15;6(3):e17971. Effects of dietary supplementation of carnosine on mitochondrial dysfunction, amyloid pathology, and cognitive deficits in 3xTg-AD mice. Corona C, Frazzini V, Silvestri E, Lattanzio R, La Sorda R, Piantelli M, Canzoniero LM, Ciavardelli D, Rizzarelli E, Sensi SL.
[20] Biofactors. 1998;8(3-4):229-34. Some new aspects of creatine kinase (CK): compartmentation, structure, function and regulation for cellular and mitochondrial bioenergetics and physiology. Wallimann T, Dolder M, Schlattner U, Eder M, Hornemann T, O'Gorman E, Rück A, Brdiczka D.
[21] J Biomed Sci. 2010 Aug 24;17 Suppl 1:S2. Physiological roles of taurine in heart and muscle. Schaffer SW, Jong CJ, Ramila KC, Azuma J.
[22] Diabetes Metab J. 2011 Jun;35(3):207-15. Mitochondrial dysfunction and insulin resistance: the contribution of dioxin-like substances. Lee HK.
[23] Cell Physiol Biochem. 2007;20(5):627-38. Aluminum-induced mitochondrial dysfunction leads to lipid accumulation in human hepatocytes: a link to obesity. Mailloux R, Lemire J, Appanna V.
[24] Toxicol Environ Chem. 2011 May;93(5-6):1251-1273. The plausibility of a role for mercury in the etiology of autism: a cellular perspective. Garrecht M, Austin DW.
[25] PLoS One. 2009;4(4):e5186. Chronic exposure to the herbicide, atrazine, causes mitochondrial dysfunction and insulin resistance. Lim S, Ahn SY, Song IC, Chung MH, Jang HC, Park KS, Lee KU, Pak YK, Lee HK.
[26] Parkinsons Dis. 2010 Dec 22;2011:951709. Toxin-induced and genetic animal models of Parkinson's disease. Hisahara S, Shimohama S.
[27] FASEB J. 2010 Dec;24(12):4989-5002. Mitochondrial dysfunction and loss of Parkinson's disease-linked proteins contribute to neurotoxicity of manganese-containing welding fumes. Sriram K, Lin GX, Jefferson AM, Roberts JR, Wirth O, Hayashi Y, Krajnak KM, Soukup JM, Ghio AJ, Reynolds SH, Castranova V, Munson AE, Antonini JM.
Osa 5: Mitokondrioiden biogeneesi
[1] Cell Metab. 2011 Feb 2;13(2):119-20. AMPK-dependent phosphorylation of ULK1 induces autophagy. Zhao M, Klionsky DJ.
[2] Proc Natl Acad Sci U S A. 2002 Dec 10;99(25):15983-7. AMP kinase is required for mitochondrial biogenesis in skeletal muscle in response to chronic energy deprivation. Zong H, Ren JM, Young LH, Pypaert M, Mu J, Birnbaum MJ, Shulman GI.
[3] Adv Physiol Educ. 2006 Dec;30(4):145-51. PGC-1alpha: a key regulator of energy metabolism. Liang H, Ward WF.
[4] Biochem J. 2009 Mar 1;418(2):261-75. AMPK and the biochemistry of exercise: implications for human health and disease. Richter EA, Ruderman NB.
[5] Am J Physiol Endocrinol Metab. 2007 Jun;292(6):E1724-39. Epub 2007 Feb 13. A high-fat, ketogenic diet induces a unique metabolic state in mice. Kennedy AR, Pissios P, Otu H, Roberson R, Xue B, Asakura K, Furukawa N, Marino FE, Liu FF, Kahn BB, Libermann TA, Maratos-Flier E. “Animals made obese on HF and transitioned to KD lost all excess body weight, improved glucose tolerance, and increased energy expenditure. Analysis of key genes showed similar changes as those seen in lean animals placed directly on KD. Additionally, AMP kinase activity was increased, with a corresponding decrease in ACC activity. These data indicate that KD induces a unique metabolic state congruous with weight loss.”
[6] Diabetes Res Clin Pract. 2008 Aug;81(2):144-9. Effect of fasting on PPARgamma and AMPK activity in adipocytes. Kajita K, Mune T, Ikeda T, Matsumoto M, Uno Y, Sugiyama C, Matsubara K, Morita H, Takemura M, Seishima M, Takeda J, Ishizuka T.
[7] Physiology (Bethesda). 2011 Aug;26(4):214-24. Calorie restriction: is AMPK a key sensor and effector? Cantó C, Auwerx J.
[8] J Physiol. 2007 Apr 15;580(Pt. 2):677-84. Upregulation of AMPK during cold exposure occurs via distinct mechanisms in brown and white adipose tissue of the mouse. Mulligan JD, Gonzalez AA, Stewart AM, Carey HV, Saupe KW.
[9] Horm Metab Res. 2012 Sep;44(9):650-5. Effect of dietary macronutrient composition on AMPK and SIRT1 expression and activity in human skeletal muscle. Draznin B, Wang C, Adochio R, Leitner JW, Cornier MA. “Our data indicate that a relative deficiency in carbohydrate intake or, albeit less likely, a relative excess of fat intake even in the absence of caloric deprivation is sufficient to activate the AMPK-SIRT 1-PGC1α energy-sensing cellular network in human skeletal muscle.”
[10] Exp Physiol. 2003 Jan;88(1):121-8. Regulation of mitochondrial biogenesis by thyroid hormone. Weitzel JM, Iwen KA, Seitz HJ.
[11] Aging (Albany NY). 2011 May;3(5):464-78. Branched-chain amino acids, mitochondrial biogenesis, and healthspan: an evolutionary perspective. Valerio A, D'Antona G, Nisoli E.
[12] Diabetes. 2009 Jul;58(7):1509-17. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Gao Z, Yin J, Zhang J, Ward RE, Martin RJ, Lefevre M, Cefalu WT, Ye J. “AMP kinase and p38 activities were elevated.”
[13] J Biol Chem. 2010 Jan 1;285(1):142-52. Pyrroloquinoline quinone stimulates mitochondrial biogenesis through cAMP response element-binding protein phosphorylation and increased PGC-1alpha expression. Chowanadisai W, Bauerly KA, Tchaparian E, Wong A, Cortopassi GA, Rucker RB.
[14] Diabetologia. 2008 Jan;51(1):165-74. R-alpha-lipoic acid and acetyl-L-carnitine complementarily promote mitochondrial biogenesis in murine 3T3-L1 adipocytes. Shen W, Liu K, Tian C, Yang L, Li X, Ren J, Packer L, Cotman CW, Liu J.
6. Yhteenveto ... Mitokondrioiden puutteellinen toiminta ... mahdollisia aiheuttajia ovat ... hapetusstressi. ... hapetusstressi vahingoittaa mitokondrion DNA:ta heikentäen mitokondrioiden toimintaa ja lisäten hapetusstressiä entisestään. ... hapetusstressiä lisäävät toimenpiteet voivat auttaa estämään, ellei jopa kumoamaan, mitokondrioiden toimintahäiriön ... EIKÖ tässä ole jokin ristiriita?
VastaaPoistaHyvä huomio, korjasin kyseisen käännösvirheen! t. Valtsu
Poista